Dramatic evidence for the localization of at least some aspect of memory came in 1953 when surgeons performed an operation called a bilateral resection of the medial temporal lobe on a patient by the name of HM. The purpose of this operation was to alleviate the intractable epilepsy which plagued him; in this matter, at least, it was a success. Unfortunately, the operation left HM with severe anterograde amnesia, which means that he could not form new memories. His retrograde amnesia, or loss of memory for events prior to the surgery, was only mild. Clearly some mechanism which helps create memories was damaged. In later years, researchers discovered that the hippocampus, which had been removed in HM's surgery, is primarily responsible for the consolidation of short-term into long-term memories. This process can take up to three years, hence HM's retrograde amnesia for events close to the surgery.
The hippocampus is not the only structure involved in memory, however. There are many different types of memory, and the hippocampus is involved in declarative, spatial, contextual, episodic, and working memories, as well as in the detection of novel stimuli. Non-declarative memories, such as motor skills, are dealt with in the neostriatum.
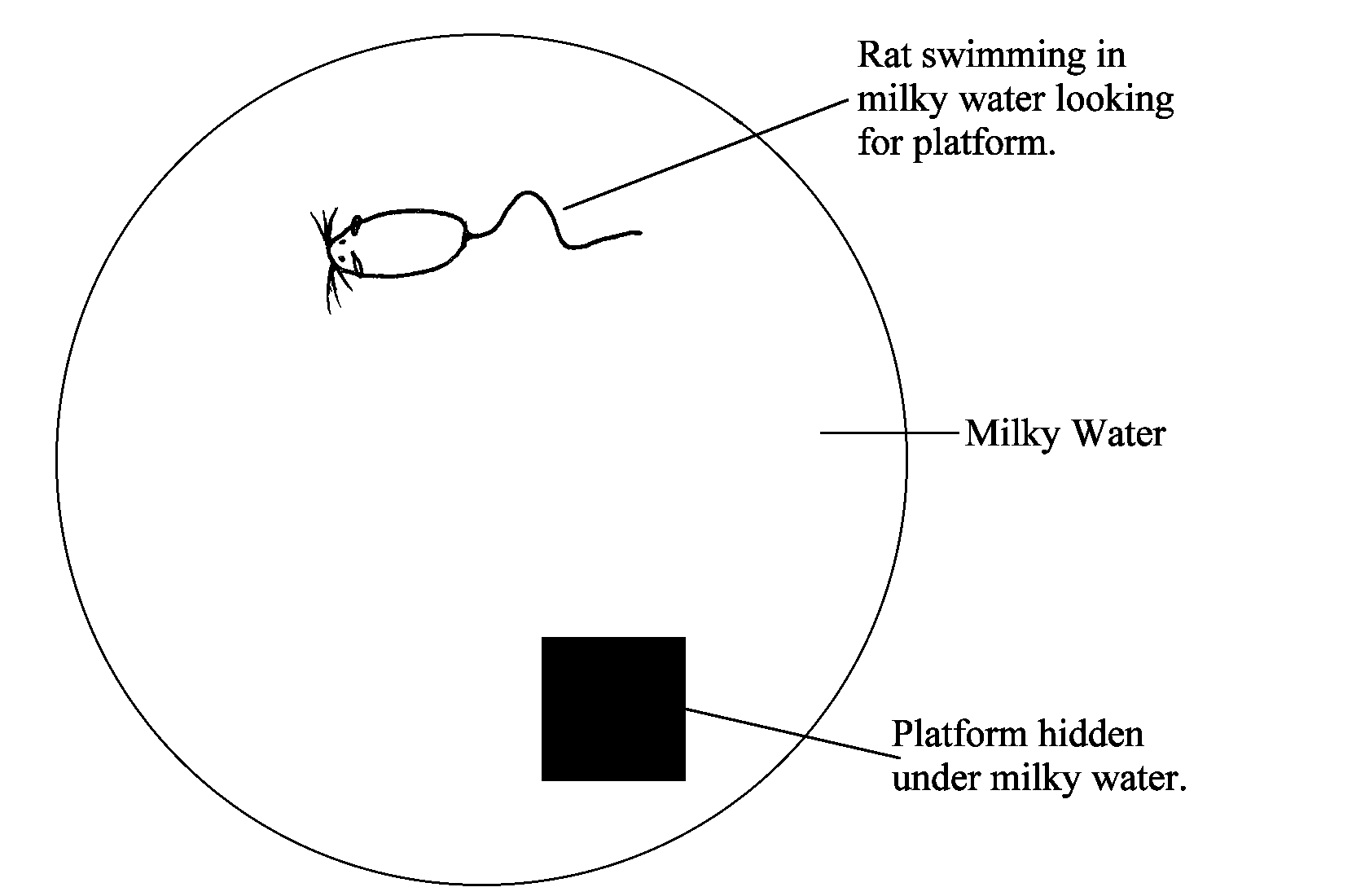
Although testing declarative memory in animals is problematic, evidence for spatial memory in them has been found. One example of the type of task used to test spatial memory is the Morris Water Maze.
In this task, a tank is filled with milky water with a platform concealed somewhere beneath the surface. When a rat is placed in the tank, he will swim around until he finds the platform. When this task is repeated many times, he learns to use cues from outside the tank and can find the platform almost immediately from any spot in the tank. He can do this because he has created a spatial map of the tank and its surroundings.
If the rat's hippocampus is lesioned after he learns the task, there will be no effect; if it is lesioned before (when he is naive), however, he is unable to learn it.
Clearly, the hippocampus is involved in creating the spatial map, but not in storing it for long periods of time. This is also evident in the fact that migratory birds have large hippocampi and as adults still experience neurogenesis in the hippocampal region.
The hippocampus is not fully developed at birth; that takes about two and one half years. An interesting effect of this is infantile amnesia--most people do not have declarative memories from their first couple years of life.
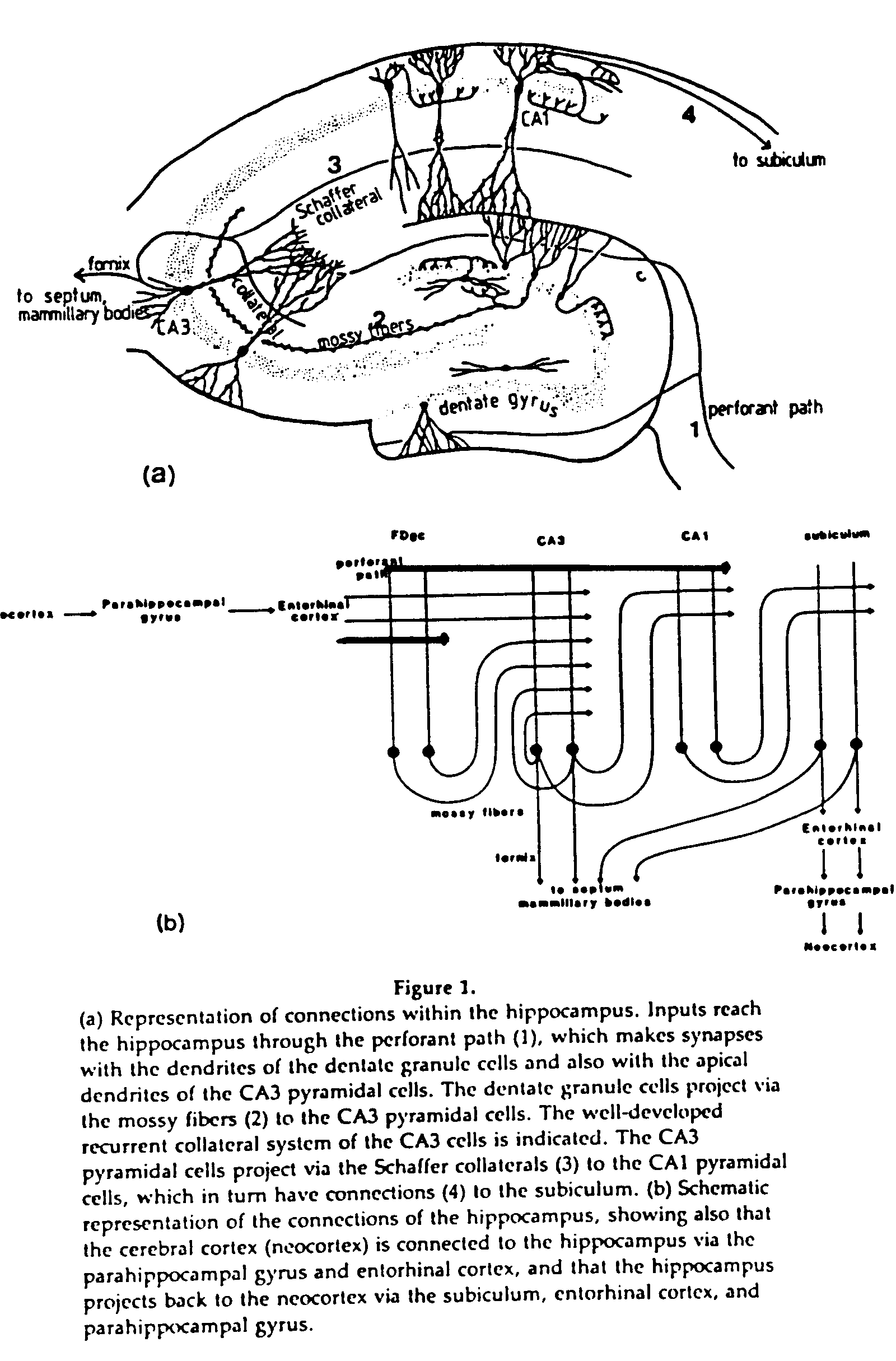
The hippocampus is a looped structure; information is funneled into it, then is sent back. It is a cylindrical structure around the thalamus. CA4 and CA2 are both quite small, and really are parts of CA3 and CA1, respectively. The hippocampal formation also includes the entorhinal cortex and a few other parts.
The main circuit is the trisynaptic pathway, which includes the perforant pathway, mossy fibers, and Schaffer collaterals. A more modern view of the connections includes staggered projections, by which each area except the dentate gyrus projects to the next two.
Back in 1949, Donald Hebb defined a type of learning which he thought occurred in cells, Hebbian Learning. It is a use-dependent mechanism which produces synaptic facilitation when pre- and post-synaptic activity is correlated.
When evidence for LTP in the hippocampus was found in 1973 by Bliss and Lomo, the process of LTP followed Hebb's principles. The mechanisms by which synaptic facilitation can occur include: an increase in the number of postsynaptic receptors or amount of neurotransmitter released, sprouting of a new terminal bouton, and structural changes which lead to less electrical resistance in the dendrite of the postsynaptic cell.
In the hippocampus, different types of structural changes occur in different areas, depending on the type of receptor involved. In the perforant pathway to the dentate gyrus, where NMDA receptors are used, they include a larger spine head, shorter neck, and a change from a convex to a concave. Between the dentate gyrus and CA3, where there are Adrenergic receptors, they include a swelling of the spine head and a wider neck.
| Location | Receptor | Before | After |
|---|---|---|---|
| DG/CA1 | NMDA |  |
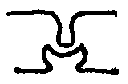 |
| CA3 | Adrenergic | |
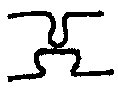 |
Gases such as NO (nitric oxide) are released by the postsynaptic cell for the presynaptic cell when activity is correlated. Gasses can diffuse over larger areas, affecting nearby synapses as well.
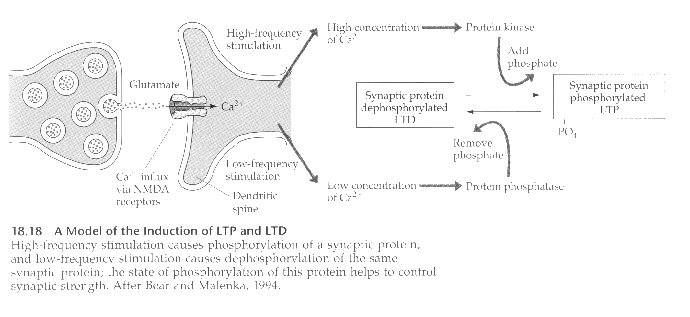
Long Term Potentiation (LTP) depends on, among other things, the presence of three types of glutamate receptors: K, Q, and NMDA. When glutamate is released by the presynaptic cell, it binds to all three types of receptors, but only the K and Q channels open to allow sodium into the cell, because the NMDA channel is blocked by a magnesium ion.
When the resulting depolarization reaches a threshold level, the magnesium block is released, opening the NMDA channel to calcium ions. Because of their size, calcium is usually prevented from entering the cell. The calcium influx then triggers a long-lasting increase in the amounts of two kinases, calimodulin kinase and protein kinase, which eventually leads to the structural changes associated with LTP.
The opposite of LTP occurs when pre- and post-synaptic activity is uncorrelated; this is called Long Term Depression (LDP). It is characterized by a decrease in synaptic efficacy.
Steps of LTP:
One interesting aspect of CA3 is its recurrent collaterals, a set of feedback fibers, which make it act as an autoassociative network. This type of network requires a set of inputs, a set of outputs, and a modifiable feedback system that connects the outputs to each other.
Computers can model autoassociative networks, which allows experimenters to discover that they have many different capabilities, including reconstructive or associative memories and acting as a novelty filter.
The reconstructive capability allows the network to complete a previously learned pattern if the input is faulty or incomplete. This allows us, for example, to recognize only a portion of a friend's face, or to associate his voice with his face. Thus it can instantiate a "content-addressable" memory to complete a learned pattern if keyed with only a portion or subset of the pattern. It also helps the network be noise resistant and fault tolerant because it can correct degraded patterns. These functions occur when the feedback connections strengthen.
If the feedback weakens, on the other hand, the network can act as a novelty filter, accentuating changed portions of a pattern.
Another interesting property of the network is that it exhibits graceful degradation. When part of the network is damaged, the rest loses its ability to recall only slowly.
The best way for an autoassociative network to learn temporal sequences of events is for the output to be fed back after a time delay. In the hippocampus, this is carried out by the staggered projections mentioned above.
The hippocampus does not store memories for long periods of time, however, but rather contains pointers which refer to information stored in locations all over the brain. Thus information from different sensory modalities can be linked into a unified whole. At some point, another mechanism must take over this job, because long term memories are often still accessible after hippocampal damage.