Part of the diencephalon, the thalamus consists of a pair of egg-shaped cell groups at the top of and continuous with the brain stem (see above figure). It is extremely important for perception, as evidenced by the fact that 98% of all sensory information is relayed through it. In fact, olfaction is the only sensory information which does not pass through the thalamus on its way to the cortex. Specialized portions of the thalamus relay the information from each sensory modality separately; association nuclei also integrate the different inputs. (See table 8.1). The thalamus also receives non-sensory input from areas important in motor control and emotion, namely the cerebellum, basal ganglia, and limbic system. Obviously, it must be performing some important function.
The prevailing hypothesis holds that the thalamus functions as a sort of "gate" or "filter," deciding what input, if any, of the multiple channels to allow through for processing. As any gate can, the thalamus can allow all, none, or some of the information through. This hypothesis was applied in a recent study of schizophrenics which found that the thalamus of a schizophrenic patient was significantly smaller than a normal patient's. The researchers proposed that schizophrenics, due to the irregularity in the thalamus, cannot "close the gate" to ignore useless or irrelevant information.
A demonstration of the gating function of the thalamus is found in EEG recordings. When a subject is awake, he shows small amplitude, high frequency (20-80 Hz) activity. When he becomes drowsy, the activity is interspersed with high amplitude, lower frequency (7-13 Hz) activity, called a rhythm. As the subject falls into progressively deeper sleep, the frequency of activity decreases while its amplitude increases. When the subject reaches REM, or "paradoxical," sleep, however, the activity resembles wakefulness.
These changes are caused by the thalamus and its interactions with its inputs and the cortex. When the thalamus allows less input through, the subject becomes more drowsy or enters deeper sleep. As he falls asleep, the gate slowly closes off progressively more channels of input. In performing this function, the thalamus also interacts with modulatory systems.
The paired, egg-shaped nuclei of the thalamus are subdivided by a fiber tract called the internal medullary lamina into several subnuclei: the dorsal medial nucleus, anteroventral nucleus, and lateral region. Of these three areas, the lateral region has shown the most marked increase in size during phylogeny. It, too, is further divided into a myriad of subnuclei, to more than forty subnuclei known.
Table 8.1:
Specific Relay Nuclei
| Inputs from | Through | Projects to |
| cochlea, auditory BN | MGN | primary auditory cortex |
| retina | LGN | primary visual cortex |
| limbic areas | AV/LD | cingulate cortex |
| spinothalamic (body) | VPL | somatosensory cortex |
| trigeminothalamic | VPM | somatosensory cortex |
| basal ganglia | VA | prefrontal, M1, premotor and supplementary motor areas |
| cerebellum | VL | prefrontal, M1, premotor and supplementary motor areas |
Association Nuclei
| Inputs from | Through | Projects to |
| superior colliculus | LP | parietal association cortex |
| amygdala, hypothalmus | DM | prefrontal association cortex |
| retina, superior colliculus, striate cortex, pretectum | Pulvinar | parietal - temporal - occipital association cortex |
Nonspecific Nuclei
| Inputs from | Through | Projects to |
| many areas, (e.g. hypothalmus and RAS) | midline and intralaminar nuclei | noncortical, sends collaterals to cortex |
The subnuclei of the thalamus are of four distinct types; classification is based on the types of input and output each nucleus handles. See Table 8.1 for a list of selected nuclei and their functions. Specific relay nuclei, as their name suggests, relay information from a specific sensory modality to its corresponding cortical area. The LGN, for example, receives input from the optic tract and sends its output to the primary visual cortex. Association nuclei receive input from specific areas and project to somewhat generalized association areas. Nonspecific nuclei both receive from and project to diffuse or nonspecific regions. Finally, subcortical nuclei create a closed loop, generating feedback to the thalamus. The reticular nucleus is of this type. Note that its inhibitory cells create an extremely fast, negative feedback loop.
In addition to the reticular nucleus feedback loop, the thalamus also is linked by a feedback circuit to the cerebral cortex. Cells in the thalamus project to layer four of the cortex; pyramidal cells in layer 6 connect back to the thalamus in much greater numbers. In fact, the ratio of feedback to projection fibers is nearly ten to one.
The neurons which make up the many thalamic nuclei are of two main types: relay cells and interneurons. Interneurons make up about 25% of the thalamic neurons. As they are inhibitory in nature, they utilize GABA.
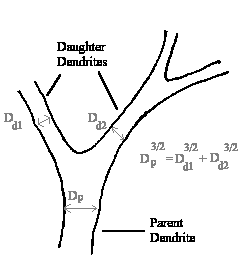
Relay cells are by far the more numerous; they account for about 75% of the total number of neurons. Their extensive dendritic arbor follows Rall's 3/2 Rule, which states that the sum of the diameters of the daughter dendrites raised to the 3/2 power equals the diameter of the parent dendrite raised to the 3/2 power. This is important because the diameter of a dendrite affects its time and length constants. Rall's Rule ensures that no inputs to a relay cell are lost; all are integrated. These inputs go to either the proximal (close to the soma) or distal (peripheral) areas of the arbor.
The different nuclei of the thalamus all preserve a separation of inputs. When input into the thalamus is topographically organized, as is visual information, the thalamus maintains this careful mapping. The LGN, for example, contains a retinotopic map and has the same cell types as exist in the retina (such as red/green cells). The information relayed to cortex is thus passed on intact. The thalamus even maintains uneven magnification present elsewhere in the brain. The inputs to the thalamus are not just sensory connections to relay cells, however.
Sensory, motor, and limbic inputs synapse onto both interneurons and the proximal dendrites of relay cells. They are, in general, excitatory, utilizing glutamate. The feedback from cortex, discussed above, is also glutamate-based and thus excitatory. It contacts interneurons and distal portions of relay cells. The feedback from the reticular nucleus, on the other hand, is inhibitory (using GABA) and synapses on proximal dendrites of relay cells; the location of its synapses suggests that it modulates sensory input. The brainstem also has its say, sending inputs to relay cells, interneurons, and the reticular nucleus from cholinergic, noradenergic, and serotonergic systems.
The gating function of the thalamus depends on these connections. Recall that an activity of 10 Hz is associated with drowsiness. If the thalamus is isolated, it will show this same 10 Hz activity. The modulatory systems in the brainstem act on the reticular nucleus, which in turn acts upon the relay cells, eventually producing 10 Hz firing by hyperpolarizing those relay cells. The result is a certain distortion of thalamic inputs.
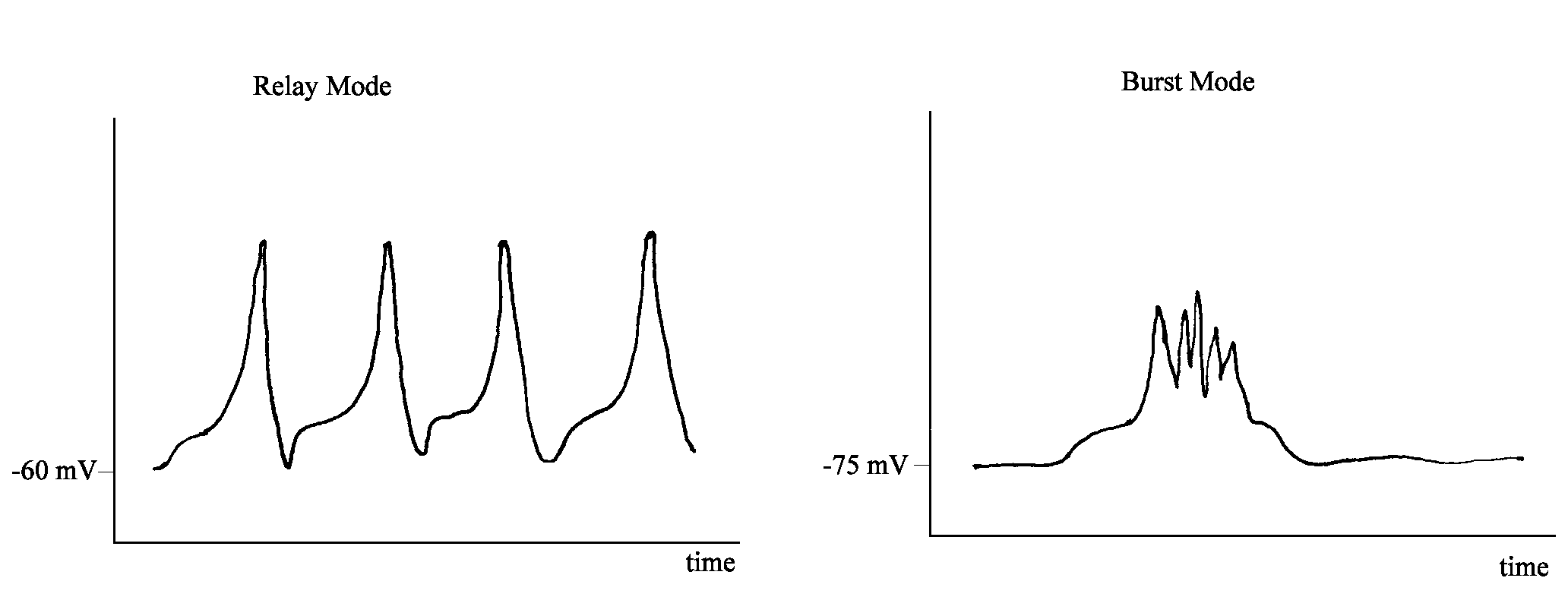
When relay cells are slightly depolarized, they fire in what is called relay mode. In this mode, there exists a linear relationship between the stimulus to a relay cell and its response. Information is thus passed on reliably to its destination. In contrast, when the relay cells are slightly hyperpolarized they fire in burst mode, a cluster of firing followed by an unresponsive period, despite that fact that the stimulus continues. Information thus is not transmitted faithfully.
One model which explains the function of the thalamus was devised by David LaBerge in 1990. He states that in order to select an item for processing from a noisy environment, there is a series of intermediate stages. They are, in order of occurrence: feature registration domain, mapping out of space, expectancy, filtering or gating, and shape identifier domain. The thalamus fulfills the function of filtering or gating.