 Figure 4.6 |
There are about 1011 to 1012 neurons (100 billion to 1 trillion) that make up the brain, each with around 103 to 104 connections. The 105 or so genes possessed by mammals could not contain sufficient information to specify all of these connections. The limits on genetic material motivates the view that epigenetic, or environmentally induced, processes activate and influence the stages of a cell's development. Our discussion of brain development (brain ontogeny) will describe the interplay of genetic and epigenetic factors. The developing embryo is made up of three layers of cells. The outer layer is called the ectoderm (Greek for "outer skin"). The middle layer is called the mesoderm ("middle skin") and the inner layer is the endoderm ("inner skin"). Human gestation takes 265 days, but, as we will see, brain development continues postnatally.
(E18-E24 or "embryonic days 18-24")
The process whereby some cells in the ectoderm of the developing embryo become transformed into the specialized tissue from which the brain and spinal cord (the CNS) develop is called neural induction. Neural induction begins when peptides are released from the mesoderm to cause cells of the ectoderm to specialize, causing it to thicken and convolute.
The central nervous system begins as the neural plate, a flat sheet of ectodermal cells on the top part of the embryo. The number of cells in the neural plate is small (close to 125,000) and this number does not change much during the formation of the neural tube. In the center of this structure, a groove appears and gradually deepens, eventually folding over itself. The plate folds into a hollow structure called the neural tube. This process begins at the midpoint of the groove and extends in both the anterior and posterior directions. Both ends of this neural tube remain temporarily open. The head end of the central canal widens to form the ventricles of the brain.
Around embryonic day 24 the anterior end closes, forming the brain. Two days later the posterior end closes, forming the spinal cord. If the rostral (front) end does not close properly, there will be improper brain formation, a condition called anancephaly (meaning no brain). If the caudal (rear) end of the neural tube does not close, spinal bifida occurs, leaving the spinal cord exposed.
In addition to the neural plate developing into the neural tube, two other structures are also seen to arise: somites and the neural crest. The somites eventually will become skeletal structures and the neural crest will form the PNS (peripheral nervous system).
(E24-E125)
Once the neural tube has been closed off, cerebrospinal fluid (which helps cushion the brain) fills the enclosed tube and cells begin to divide at a furious rate (250,000 cells /minute) forming several layers. At the end of the 4th week, the three primary parts of the brain can be seen at the head or anterior part of the neural tube. These are the forebrain, midbrain, and hindbrain. At the 5th week of development the forebrain (prosencephalon) develops into the telencephalon (cerebral cortex, basal ganglia, and the limbic system) and the diencephalon (thalamus and hypothalamus). The midbrain or mesencephalon develops into the superior and inferior colliculi. The hindbrain (rhombencephalon) develops into the mesencephalon (cerebellum and pons) and myelencephalon (medulla).
There are four stages of cell division that must occur in order continue neurogenesis, the production of nerve cells. 1) In the G1 phase, cells move from the ventricular zone to the marginal zone of the neural tube. 2) In the S phase, the cells which are genetically programmed to divide return to the ventricular zone. 3) These cells copy their DNA in the G2 phase to prepare for mitosis (cell division) in the 4) M phase.
Then at a critical period, around E40, some cells in all regions of the neural tube lose their capacity to divide or reproduce. This date is referred to as a cell's "birthday". Only neurons lose their ability to divide. Glial cells can keep dividing. Neurons carry electrical impulses and glial cells provide supportive services (i.e. hold neurons in place, control supplies of various chemicals needed for neural communication, insulate neurons to enhance signal transmission, and remove waste products).
(E40-E160)
Once a cell's "birthday" occurs, migration can begin. Migration involves the movement of a cell to its final destination. Neurons attach to radial glial cells, specialized glial cells, and climb along them till they reach their destination. This is a slow way of moving (a tenth of a millimeter per day). Radial glial cells act as guides for the neurons - that is they guide them to where they are supposed to end up. There are special molecules on the cell's surface that allow it to recognize cells of the same kind called CAM's (cell adhesion molecules). One special feature of cell migration in the developing nervous system is that most regions of the brain not only adhere to one another but also adopt some preferential orientation. Cells that get lost on their way and end up where they are not supposed to be constitute ectopias. These errors are typically eliminated during the later stages but in some cases lead to severe brain disorders. Ectopias are not caused by having extra cells in some areas of the brain, but by insufficient cells in other parts. Problems with radial glial cell trafficking may lead to schizophrenia and dyslexia.
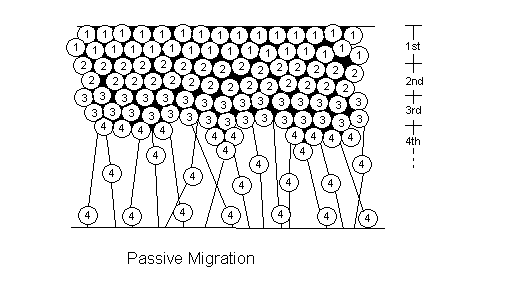
There are two types of cell migration: active and passive (See figure 4.6). In non-laminated (nonlayered) areas of the brain (i.e. basal ganglia, thalamus, hypothalamus), the neurons formed first in the ventricular zone end up at the top at the marginal zone. This is passive migration: the cells born first have been pushed up by the cells growing underneath them. Active migration, which only occurs in laminated (layered) areas (i.e. cortex, hippocampus, colliculi), differs in that cells formed last are the ones ending up nearest to the marginal zone. Newly created cells actively climb to the top of the mountain using radial glial cells as their rope (See figure 4.7).
Figure 4.6 |
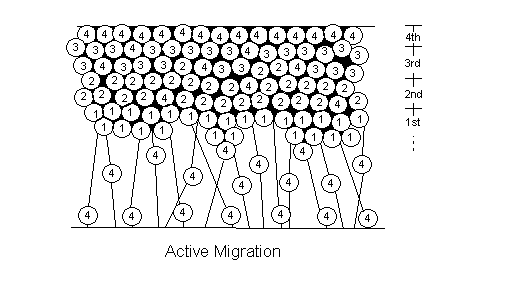 Figure 4.7 |
There are three phases to migration: 1) the transition phase. where the neuron becomes attached to the glial cell; 2) the locomotory phase, during which the neuron moves at the rate of 100 micrometers per day: and 3) the recognition phase, which is when the neuron recognizes its destination and stops migrating.
(E125-postnatal)
The stage of differentiation, which begins once the cell has found its destination, involves axon growth and dendritic arborization, as well as the adoption of a way of communicating with other cells. This constitutes cell specialization. Most cells in the adult brain are multipolar, having processes radiating in all directions. There are some bipolar cells, with only two processes and a few unipolar cells, with only one process. As a rule, neurons have one very long process called an axon, which carries the output signal from the cell, and many dendrites which receive the input from other cells. During normal development of the brain most neurons are subject to a variety of environmental influences that may modify their form. Cell to cell communication and interaction is critical at this point.
There are several ways in which cells communicate: one is through electrical impulses called action potentials. Most neurons do not physically connect with each other, rather there are small gaps between neurons. These gaps are called synapses (Greek for "junction"). Electrical potentials flow across a cell's axon to the end of the neuron where they release chemicals. These chemicals, called neurotransmitters, swim across the synapse and start the electrical impulse on the next neuron. In some cases neurons are actually connected and there is no discontinuity in the electrical impulses. There is another way of communication through what are called gap junctions. This stage of differentiation is where it is determined how communication between cells will occur. Under the influence of certain environmental factors cells can change their method of electrical impulse. They can change the type of neurotransmitter they use to communicate with other cells, as well as a host of other things that while initially predetermined can be dynamically altered. This underlies the fact that the brain is not a static organ, rather it is a dynamic, living thing, and nature plays important roles in its development. Thus, this stage possesses both genetic and epigenetic aspects: experience and/or the cell's environment may induce various responses of an organizational nature.
There are approximately 100 billion cells in the brain and perhaps the most important stage during development and the one thing that is very poorly understood is how these cells connect with one another, and the pattern of those connections. We know that each of the neuron's axons and dendrites have what are called growth cones on the end (See figure 4.8). Growth cones help guide cells to their target location, where the cell may then be influenced by neighboring cells in its process of specialization. The growth cones have filopodia which are proteins that extend and retract, that lead the cells. They have sensors for sensing and sampling the environment. It is believed that molecules found on the surface of these growth cones act to recognize specific chemicals released by target cells. These chemicals form a gradient. This hypothesis, known as "chemotrophic guidance of axons", is based on a chemical attraction for growing axons.
A cell forms its connections by using two strategies to reach its target: long distance zeroing in on an area and finding local address within that area. The long distance strategy is mediated by performed pathways, which consist of radial glial scaffolding and pioneer neurons. It can also be mediated by distributed positional cues (guide posts). A third way that the long distance strategy is mediated is by chemotropism, which is diffusible chemical gradients that attract or repel (this refers back to growth cones) Local address connections are made through a matching stage and a sorting and adjustment stage. The matching stage shows low precision and high exuberancy. The sorting and adjustment stage shows a stabilization of some connections and the removal of others.
| "... one could say that the growth cone is a sort of battering
ram, endowed with an : exquisite chemical sensitivity and advancing by- rapid ameboid
movements that allow it to swerve around and: surmount obstacles in its path, thus forcing
a passage between; cells until it reaches its destination" Ramón y Cajal, 1899 |
As these synapses are forming, there are two types of connection pruning to consider cell death and elimination. Neurons at the early stages of development develop far more branches than are found in the mature state. There is an over production of cells and connections. As the organism grows, however, only a subset of these connections is maintained, while the remaining connections are eliminated or retracted. For these reasons, cell death, also called apoptosis, and elimination are thought of as the final brain developmental stages, which may in fact continue for the remainder of our lives.
The process of synapse rearrangement, also referred to as synapse remodeling, is named such because some synapses are lost and new ones are also formed. This process usually takes place after cell death. Both elimination and retraction are both part of synapse rearrangement.
There are several factors that play in the process of elimination. One is the activity of the cell. The more active a cell is, the less likely it will be eliminated. Second, competition from other cells plays an important part. It is believed that neurons compete with other neurons for a variety of things, including areas to enervate and chemicals that help nourish and maintain them (i.e. growth factors).
In a region where there is a lot of competition, more cells will be eliminated than where there is little competition. This process of competition and elimination has been called Neural Darwinism.
Two more terms should be introduced here that will help us distinguish between cell death and elimination processes. Convergent exuberancy describes the situation in which many more source neurons than are necessary or used have processes extended to a target neuron. Some of these source neurons will die off so that they won't use up chemicals needed by the more important source neurons. Divergent exuberancy, in contrast, refers to the case where many more processes that are necessary diverge from a single source neuron, and some of these unneeded processes get "eliminated" (decompose or retract).
It should be noted that while the number of cells is a maximum at the time of birth, the net number of connections increases after that point since learning requires the formation of new synapses. How connections are made is the basis of Hebbian Learning. Donald Hebb found there to be a connection between positive correlations of pre- and post-synaptic cells and learning. This positive correlation will increase the efficacy of the synapse, known as long term potentiation (LTP). The inverse is also true: if there is no correlation (negative correlation), then long term depression (LTD) occurs and this is a weakening of the synapse.
Cell death and synapse rearrangement can be thought of as phenomena in which epigenetic factors (experience) induce genetically provided mechanisms (cell or process degradation).