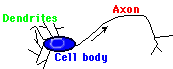
A nerve can be divided into two functional parts:
| Dendrites are short, thin, highly branched, and are passive conductors of electrical impulses; axons may be very long (e.g., the spinal chord), generally have few branches (collateral axons), and actively transmit nerve impulses. This diagram is a highly simplified picture of a simple nerve; in reality there would be many dendrites and thousands of synaptic inputs into them. |
|
Mechanism: Nerves generate electricity chemically. Ions (electrically charged atoms or molecules) are actively transported across the cell membrane -- from inside the cell (cytoplasm) to outside (interstitial fluid), and vice versa. By selectively "pumping" different ions, the cell membrane acts to make the inside of the cell negatively charged relative to the outside. The major ions are sodium, potassium, chlorine, and calcium (Na, K, Cl, & Ca); at rest, the difference generated is usually about -60 to -90 millivolts. The membrane is polarized.
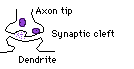 | Now the entering impulse. Molecules of neurotransmitter are released from the synaptic vessicles in which they are stored, and diffuse into the synaptic cleft. This being very narrow, many/most contact the other side where they bind with specific receptor molecules on the postsynaptic cell membrane. |
This is analogous to a lock and key mechanism, apparently even to the critical factor being the match between the physical shapes of the transmitter and receptor molecules. When the transmitter "clicks into place," it causes the receptor molecule to change shape, and this sets in motion a set of physical/chemical interactions in the membrane of the postsynaptic cell that result in a sudden increase in the permeability of the membrane near the receptor to specific ions. When this happens, positive ions (mainly Na+) are pulled into the cell by the negatively charged cytoplasm (osmotic pressures and other ions are involved and complicate this simplistic picture).
The flow of ions across the membrane is an electrical event, and if enough transmitter has been received this depolarization creates an electrical impulse that is carried along the dendrite to the cell body. The trick here is that the ion flow itself causes neighboring "gates" in the membrane to "open" to Na+ and so the process is self-reinforcing. The nerve impulse itself is simply a self-propagating electrical event that moves the length of the axon; the current crossing the membrane causes the membrane just ahead to let in Na+ (current) which of course affects the next bit of membrane.
This is the basic idea behind the impulse. I haven't distinguished between passive (dendritic) and active (axonal) transmission; the observed difference is that dendritic impulses lose strength over distances, whereas active impulses -- action potentials -- do not.
Because it is self-maintaining, the axonal impulse is all-or-none: varying the stimulus intensity (i.e., dumping few or many vessicles, or applying a mild or strong electrical current) can have one of only two outcomes -- the impulse is triggered or it's not. Think about what this lack of individually graded response means for information processing.
When the impulse reaches the end of the axon, the changes it elicits in the cell membrane cause vessicles to be released, dumping neurotransmitter molecules, and we're back to the beginning.
Going from how a neuron sends signals to how information is acquired, stored, and retrieved is a bit of a jump even for this course. If you want to tackle it, look into a major in cogsci or physiological psych. Presumably at least part of the story lies in wiring changes; nerves grow dendrites and make connections as a function of experience/stimulation.
Loose ends:
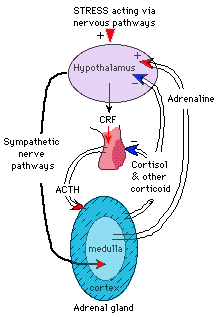 | A good example of a system that uses nerves and hormones is the stress response. A stressful event (a gun going off unexpectedly, say, or an announcement of a pop quizz...) is perceived, and the appropriate nerves in the hypothalamus are stimulated. Some of these have direct axonal connections to the adrenal medulla (the adrenal glands are next to your kidneys; the inside is the medula and the outer layer is the cortex). When these fire, medullary cells release adrenalin, which causes the rush you feel as you mobilize your body -- it acts as a stimulant to get you out of harm's way. Other nerves in the hypothalamus release corticotropin releasing factor (CRF) which is carried to the anterior pituitary where it stimulates the release of ACTH. The ACTH is carried in the blood to the adrenal cortex, where it stimulates the release of cortisol -- a more low-keyed, but longer-lasting, mobilizer of the body's energy. As you can imagine, the hormone system takes longer to start up; but once going, it keeps working longer. A rule of thumb -- nerves are better for fast, acute responses; hormones for slower, chronic ones
(but note that slow is only relative; it takes only a few seconds for ACTH to start releasing cortisol).
Note the pinkish gland in the center (figuratively as well as literally) is the pituitary; the anterior pituitary (lighter color) is not innervated from hypothalamus; posterior pituitary receives direct nervous inputs and secretes e.g. oxytocin. "+" and red arrows and "-" and blue arrows signs indicate positive & negative feedback. (After Clegg & Clegg [1969] Hormones, Cells and Organisms. Stanford University Press.) |
Feedback control: Although feedback "loops" are involved in the function of both endocrine and nervous systems, hormonal loops are both simpler and probably more important to the total system than are nerve loops (imagine a "loop" involving a hundred thousand cells and several million synapses -- the principle is similar, but the actual description is way beyond us). This discussion therefore focuses only on hormonal systems.
The principle is simple: hormone secreted by a cell may act, directly or indirectly, to increase (positive feedback) or to decrease (negative feedback, the more common type) the output of that hormone. For example -- cell A receives a nervous or chemical message to secrete hormone X, which it begins to do. Cell A has receptors in its membrane that, when bound with X, alter the enzyme reaction such that the secretion of X is halted. Clearly, as the blood level of X rises it will "turn itself off" -- direct negative feedback. Complexity is introduced into the system by the potential for indirect feedback (e.g., X causes cell B to release Y which, among other things, causes neuron C to fire, causing D to release Z which turns off cell A...) and by systems or inputs controlling the loops and that sort of thing.
E.g., the stress system mentioned above. Adrenalin released from the adrenal medulla stimulates hypothalamic cells that secrete CRF, which are already active due to nerve inputs. This results in a high CRF (--> ACTH --> cortisol) output. But cortisol has a negative effect on the production of CRF and ACTH; as cortisol builds up and nerves slow their repsonse to the stressor (you got away, or the lion wasn't hungry, etc.) a crossover is reached where the negative feedback outweighs the stimulation and the stress response turns itself off.
| Vocabulary: There are almost as many terms as chemicals floating around. Some guidlines: "H," as in LH, means hormone. "F" in a similar context means "factor," which is what it's called when everybody knows it's there but nobody knows exactly what it is yet. Thus, you may see both LHRF and LHRH; LHRF is just the older term. "Epinepherine" is just British for adrenalin. | 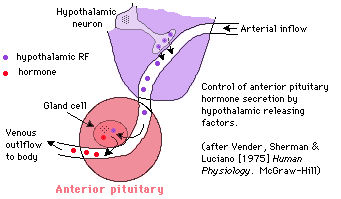 |
Many hormones can be kept straight by knowing what the names mean. FSH is follicle stimulating hormone; egg cells are contained in follicles (look it up) within the ovary that must develop before ovulation. So -- what does FSH do? Similarly, pro- is a prefix meaning "for" or "in favor of;" you know what lactation and gestation mean, so what do prolactin and progesterone do? And again, it's no surprise that LH (luteinizing hormone) causes ovulation and stimulates the resulting corpus luteum (itself an endocrine organ). Adrenalin (adrenal), cortisol (adrenal cortex), and thyroxine (thyroid) all tell you where they're produced.
Releasing hormones (RF or RH) are produced by neurosecretory cells -- essentially nerves that "synapse" into the circulatory system instead of other cells -- and cause the release of other hormones. So far, all RHs are hypothalamic. Tropic (or trophic) hormones are released by one endocrine gland and affect a different one. Now -- since the chorion is a membrane around the embryo, it's hopefully no surprise that human chorionic gonadotropin (HCG) is released by the chorion and affects hormone secretion by the gonads of a pregnant woman. And what about ACTH -- adreno cortico tropic hormone?
Origins of sex (and hormones):
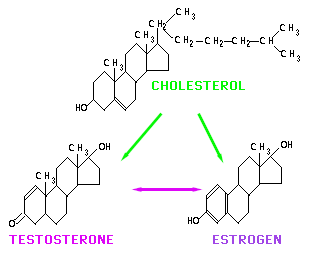 |
People sometimes view "male" and "female" as opposites. This is very much an exageration. One might think in terms of a broad gradation from extremely "masculine" to very "feminine;" people near the "masculine" end tending to be males and people near the "feminine" end more often being female. In the middle it can get hard to differentiate the sexes in terms of behaviors that aren't actually related to sex and parenting. Research on sex and gender is a fascinating subfield in anthropology, one that can be approached from either the cultural or the biological side. For now: on the next page are schematics of the chemical structures for the two major "sex hormones" and their precursor, cholesterol. Don't sweat the details -- you don't need to be a rocket scientist to see the similarities. In fact, one of the steps in going from cholesterol to estradiol 17 b (the primary estrogen in humans) is testosterone.
|
Dietary cholesterol (above) provides the basic structure for the synthesis of testosterone (left) and estradiol 17 b (right). Both are found in the blood of both men and women, though generally in very different amounts.
The problem of the mechanism of hormone action is difficult. ... The hormone, very often after binding to a specific tissue or tissue component, might then: